
根系吸收的氮主要是无机态氮,即铵态氮和硝态氮,也可吸收一部分有机态氮,如尿素。
氮是蛋白质、核酸、磷脂的主要成分,而这三者又是原生质、细胞核和生物膜的重要组成部分,它们在生命活动中占有特殊作用。因此,氮被称为生命的元素。
酶以及许多辅酶和辅基如NAD+、NADP+、FAD等的构成也都有氮参与。氮还是某些植物激素如生长素和细胞分裂素、维生素如B1、B2、B6、PP等的成分,它们对生命活动起重要的调节作用。
氮是叶绿素的成分,与光合作用有密切关系。由于氮具有上述功能,所以氮的多寡会直接影响细胞的分裂和生长。
当氮肥供应充足时,植株枝叶繁茂,躯体高大,分蘖(分枝)能力强,籽粒中含蛋白质高。植物必需元素中,除碳、氢、氧外,氮的需要量最大,因此,在农业生产中特别注意氮肥的供应。常用的人粪尿、尿素、硝酸铵、硫酸铵、碳酸氢铵等肥料,主要是供给氮素营养。
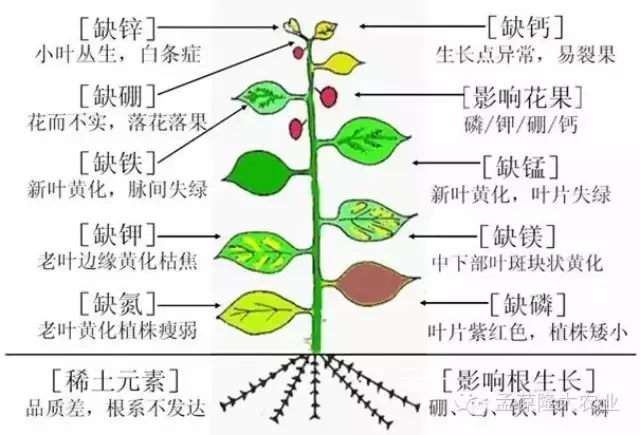
缺氮时,蛋白质、核酸、磷脂等物质的合成受阻,植物生长矮小,分枝、分蘖很少,叶片小而薄,花果少且易脱落;缺氮还会影响叶绿素的合成,使枝叶变黄,叶片早衰甚至干枯,从而导致产量降低。因为植物体内氮的移动性大,老叶中的氮化物分解后可运到幼嫩组织中去重复利用,所以缺氮时叶片发黄,由下部叶片开始逐渐向上,这是缺氮症状的显著特点。
氮过多时,叶片大而深绿,柔软披散,植株徒长。另外,氮素过多时,植株体内含糖量相对不足,茎秆中的机械组织不发达,易造成倒伏和被病虫害侵害。
磷主要以H2PO4-或HPO42-的形式被植物吸收。吸收这两种形式的多少取决于土壤pH,pH<7时,H2PO4-居多;pH>7时,HPO42-较多。
当磷进入根系或经木质部运到枝叶后,大部分转变为有机物质如糖磷脂、核苷酸、核酸、磷脂等,有一部分仍以无机磷形式存在。植物体中磷的分布不均匀,根、茎的生长点较多,嫩叶比老叶多,果实、种子中也较丰富。
磷是核酸、核蛋白和磷脂的主要成分,它与蛋白质合成、细胞分裂、细胞生长有密切关系;磷是许多辅酶如NAD+、NADP+等的成分,它们参与了光合、呼吸过程;磷是AMP、ADP和ATP的成分;磷还参与碳水化合物的代谢和运输,如在光合作用和呼吸作用过程中,糖的合成、转化、降解大多是在磷酸化后才起反应的;磷对氮代谢也有重要作用,如硝酸还原有NAD+和FAD的参与,而磷酸吡哆醛和磷酸吡哆胺则参与氨基酸的转化;磷与脂肪转化也有关系,脂肪代谢需要NADPH、ATP、CoA(辅酶)和NAD+的参与。
由于磷参与多种代谢过程,而且在生命活动最旺盛的分生组织中含量很高,因此施磷对分蘖、分枝以及根系生长都有良好作用。由于磷促进碳水化合物的合成、转化和运输,对种子、块根、块茎的生长有利,故马铃薯、甘薯和禾谷类作物施磷后有明显的增产效果。由于磷与氮有密切关系,所以缺氮时,磷肥的效果就不能充分发挥。只有氮磷配合施用,才能充分发挥磷肥效果。总之,磷对植物生长发育有很大的作用,是仅次于氮的第二个重要元素。
缺磷会影响细胞分裂,使分蘖分枝减少,幼芽、幼叶生长停滞,茎、根纤细,植株矮小,花果脱落,成熟延迟;缺磷时,蛋白质合成下降,糖的运输受阻,从而使营养器官中糖的含量相对提高,这有利于花青素的形成,故缺磷时叶子呈现不正常的暗绿色或紫红色,这是缺磷的病症。
磷在体内易移动,也能重复利用,缺磷时老叶中的磷能大部分转移到正在生长的幼嫩组织中去。因此,缺磷的症状首先在下部老叶出现,并逐渐向上发展。
磷肥过多时,叶上又会出现小焦斑,系磷酸钙沉淀所致;磷过多还会阻碍植物对硅的吸收,易招致水稻感病。水溶性磷酸盐还可与土壤中的锌结合,减少锌的有效性,故磷过多易引起缺锌病。
钾在土壤中以KCl、K2SO4等盐类形式存在,在水中解离成K+而被根系吸收。在植物体内钾呈离子状态。钾主要集中在生命活动最旺盛的部位,如生长点,形成层,幼叶等。
钾在细胞内可作为60多种酶的活化剂,如丙酮酸激酶、果糖激酶、苹果酸脱氢酶、琥珀酸脱氢酶、淀粉合成酶、琥珀酰CoA合成酶、谷胱甘肽合成酶等。因此钾在碳水化合物代谢、呼吸作用及蛋白质代谢中起重要作用。钾能促进蛋白质的合成,钾充足时,形成的蛋白质较多,从而使可溶性氮减少。钾与蛋白质在植物体中的分布是一致的,例如在生长点、形成层等蛋白质丰富的部位,钾离子含量也较高。富含蛋白质的豆科植物的籽粒中钾的含量比禾本科植物高。
钾与糖类的合成有关。大麦和豌豆幼苗缺钾时,淀粉和蔗糖合成缓慢,从而导致单糖大量积累;而钾肥充足时,蔗糖、淀粉、纤维素和木质素含量较高,葡萄糖积累则较少。钾也能促进糖类运输到贮藏器官中,所以在富含糖类的贮藏器官(如马铃薯块茎、甜菜根和淀粉种子)中钾含量较多。此外,韧皮部汁液中含有较高浓度的K+,约占韧皮部阳离子总量的80%。从而推测K+对韧皮部运输也有作用。
K+是构成细胞渗透势的重要成分。在根内K+从薄壁细胞转运至导管,从而降低了导管中的水势,使水分能从根系表面转运到木质部中去;K+对气孔开放有直接作用见表2-5,离子态的钾,有使原生质胶体膨胀的作用,故施钾肥能提高作物的抗旱性。
缺钾时,植株茎杆柔弱,易倒伏,抗旱、抗寒性降低,叶片失水,蛋白质、叶绿素破坏,叶色变黄而逐渐坏死。缺钾有时也会出现叶缘焦枯,生长缓慢的现象,由于叶中部生长仍较快,所以整个叶子会形成杯状弯曲,或发生皱缩。钾也是易移动可被重复利用的元素,故缺素病症首先出现在下部老叶。
N、P、K是植物需要量很大,且土壤易缺乏的元素,故称它们为“肥料三要素”。

植物从土壤中吸收CaCl2、CaSO4等盐类中的钙离子。钙离子进入植物体后一部分仍以离子状态存在,一部分形成难溶的盐(如草酸钙),还有一部分与有机物(如植酸、果胶酸、蛋白质)相结合。钙在植物体内主要分布在老叶或其它老组织中。
钙是植物细胞壁胞间层中果胶酸钙的成分,因此,缺钙时,细胞分裂不能进行或不能完成,而形成多核细胞。钙离子能作为磷脂中的磷酸与蛋白质的羧基间联结的桥梁,具有稳定膜结构的作用。
钙对植物抗病有一定作用。据报道,至少有40多种水果和蔬菜的生理病害是因低钙引起的。苹果果实的疮痂病会使果皮受到伤害,但如果供钙充足,则易形成愈伤组织。钙可与植物体内的草酸形成草酸钙结晶,消除过量草酸对植物(特别是一些含酸量高的肉质植物)的毒害。钙也是一些酶的活化剂,如由ATP水解酶、磷脂水解酶等酶催化的反应都需要钙离子的参与。
植物细胞质中存在多种与Ca2+有特殊结合能力的钙结合蛋白(calcium binding proteins,CBP),其中在细胞中分布最多的是钙调素(Calmodulin,CaM)。Ca2+与CaM结合形成Ca2+—CaM复合体,它在植物体内具有信使功能,能把胞外信息转变为胞内信息,用以启动、调整或制止胞内某些生理生化过程。
缺钙初期顶芽、幼叶呈淡绿色,继而叶尖出现典型的钩状,随后坏死。钙是难移动,不易被重复利用的元素,故缺素症状首先表现在上部幼茎幼叶上,如大白菜缺钙时心叶呈褐色。
镁以离子状态进入植物体,它在体内一部分形成有机化合物,一部分仍以离子状态存在。
镁是叶绿素的成分,又是RuBP羧化酶、5-磷酸核酮糖激酶等酶的活化剂,对光合作用有重要作用;镁又是葡萄糖激酶、果糖激酶、丙酮酸激酶、乙酰CoA合成酶、异柠檬酸脱氢酶、α酮戊二酸脱氢酶、苹果酸合成酶、谷氨酰半胱氨酸合成酶、琥珀酰辅酶A合成酶等酶的活化剂,因而镁与碳水化合物的转化和降解以及氮代谢有关。镁还是核糖核酸聚合酶的活化剂,DNA和RNA的合成以及蛋白质合成中氨基酸的活化过程都需镁的参加。具有合成蛋白质能力的核糖体是由许多亚单位组成的,而镁能使这些亚单位结合形成稳定的结构。如果镁的浓度过低或用EDTA(乙二胺四乙酸)除去镁,则核糖体解体,破裂为许多亚单位,蛋白质的合成能力丧失。因此镁在核酸和蛋白质代谢中也起着重要作用。
缺镁最明显的病症是叶片贫绿,其特点是首先从下部叶片开始,往往是叶肉变黄而叶脉仍保持绿色,这是与缺氮病症的主要区别。严重缺镁时可引起叶片的早衰与脱落。
硫主要以SO42-形式被植物吸收。硫酸根离子进入植物体后,一部分仍保持不变,而大部分则被还原成S,进而同化为含硫氨基酸,如胱氨酸,半胱氨酸和蛋氨酸。这些氨基酸是蛋白质的组成成分,所以硫也是原生质的构成元素。辅酶A和硫胺素、生物素等维生素也含有硫,且辅酶A中的硫氢基(-SH)具有固定能量的作用。硫还是硫氧还蛋白、铁硫蛋白与固氮酶的组分,因而硫在光合、固氮等反应中起重要作用。另外,蛋白质中含硫氨基酸间的-SH基与-S-S-可互相转变,这不仅可调节植物体内的氧化还原反应,而且还具有稳定蛋白质空间结构的作用。由此可见,硫的生理作用是很广泛的。
硫不易移动,缺乏时一般在幼叶表现缺绿症状,且新叶均衡失绿,呈黄白色并易脱落。缺硫情况在农业上很少遇到,因为土壤中有足够的硫满足植物需要。

铁主要以Fe2+的螯合物被吸收。铁进入植物体内就处于被固定状态而不易移动。铁是许多酶的辅基,如细胞色素、细胞色素氧化酶、过氧化物酶和过氧化氢酶等。在这些酶中铁可以发生Fe3++e-==Fe2+的变化,它在呼吸电子传递中起重要作用。细胞色素也是光合电子传递链中的成员(Cytf和Cytb559、Cytb563),光合链中的铁硫蛋白和铁氧还蛋白都是含铁蛋白,它们都参与了光合作用中的电子传递。
铁是合成叶绿素所必需的,其具体机制虽不清楚,但催化叶绿素合成的酶中有两三个酶的活性表达需要Fe2+。近年来发现,铁对叶绿体构造的影响比对叶绿素合成的影响更大,如眼藻虫(Euglena)缺铁时,在叶绿素分解的同时叶绿体也解体。另外,豆科植物根瘤菌中的血红蛋白也含铁蛋白,因而它还与固氮有关。
铁是不易重复利用的元素,因而缺铁最明显的症状是幼芽幼叶缺绿发黄,甚至变为黄白色,而下部叶片仍为绿色。土壤中含铁较多,一般情况下植物不缺铁。但在碱性土或石灰质土壤中,铁易形成不溶性的化合物而使植物缺铁。
在通气良好的土壤中,铜多以Cu2+的形式被吸收,而在潮湿缺氧的土壤中,则多以Cu+的形式被吸收。Cu2+以与土壤中的几种化合物形成螯合物的形式接近根系表面。
铜为多酚氧化酶、抗坏血酸氧化酶、漆酶的成分,在呼吸的氧化还原中起重要作用。铜也是质蓝素的成分,它参与光合电子传递,故对光合有重要作用。铜还有提高马铃薯抗晚疫病的能力,所以喷硫酸铜对防治该病有良好效果。植物缺铜时,叶片生长缓慢,呈现蓝绿色,幼叶缺绿,随之出现枯斑,最后死亡脱落。另外,缺铜会导致叶片栅栏组织退化,气孔下面形成空腔,使植株即使在水分供应充足时也会因蒸腾过度而发生萎蔫。
硼以硼酸(H3BO3)的形式被植物吸收。高等植物体内硼的含量较少,约在2~95mg•L-1范围内。植株各器官间硼的含量以花最高,花中又以柱头和子房为高。硼与花粉形成、花粉管萌发和受精有密切关系。缺硼时花药花丝萎缩,花粉母细胞不能向四分体分化。
用14C标记的蔗糖试验证明,硼能参与糖的运转与代谢。硼能提高尿苷二磷酸葡萄糖焦磷酸化酶的活性,故能促进蔗糖的合成。尿苷二磷酸葡萄糖(UDPG)不仅可参与蔗糖的生物合成,而且在合成果胶等多种糖类物质中也起重要作用。
硼还能促进植物根系发育,特别对豆科植物根瘤的形成影响较大,因为硼能影响碳水化合物的运输,从而影响根对根瘤菌碳水化合物的供应。因此,缺硼可阻碍根瘤形成,降低豆科植物的固氮能力。此外,用14C—半氨基酸的标记试验发现,缺硼时氨基酸很少参入到蛋白质中去,这说明缺硼对蛋白质合成也有一定影响。
不同植物对硼的需要量不同,油菜、花椰菜、萝卜、苹果、葡萄等需硼较多,需注意充分供给;棉花、烟草、甘薯、花生、桃、梨等需量中等,要防止缺硼;水稻、大麦、小麦、玉米、大豆、柑橘等需硼较少,若发现这些作物出现缺硼症状,说明土壤缺硼已相当严重,应及时补给。
缺硼时,受精不良,籽粒减少。小麦出现的“花而不实”和棉花上出现的“蕾而不花”等现象也都是因为缺硼的缘故。
缺硼时根尖、茎尖的生长点停止生长,侧根侧芽大量发生,其后侧根侧芽的生长点又死亡,而形成簇生状。甜菜的干腐病、花椰菜的褐腐病、马铃薯的卷叶病和苹果的缩果病等都是缺硼所致。

锌以Zn2+形式被植物吸收。锌是合成生长素前体—色氨酸的必需元素,因锌是色氨酸合成酶的必要成分,缺锌时就不能将吲哚和丝氨酸合成色氨酸,因而不能合成生长素(吲哚乙酸),从而导致植物生长受阻,出现通常所说的“小叶病”,如苹果、桃、梨等果树缺锌时叶片小而脆,且丛生在一起,叶上还出现黄色斑点。北方果园在春季易出现此病。
锌是碳酸酐酶(carbonic anhydrase,CA)的成分,此酶催化CO2+H2O=H2CO3的反应。由于植物吸收和排除CO2通常都先溶于水,故缺锌时呼吸和光合均会受到影响。锌也是谷氨酸脱氢酶及羧肽酶的组成成分,因此它在氮代谢中也起一定作用。
植物缺锌较严重时会出现很多症状,主要是叶片褪绿黄白化,叶形显著变小,茎节间缩短,常发生小叶丛生,称为“小叶病”、“簇叶病”等果实小、变形,核果桨果的果肉有紫斑,生长缓慢,植株矮。
树缺锌常出现“小叶病”;
玉米苗期缺锌出现“花白苗”;
水稻缺锌引起“火烧苗”;
小麦缺锌节间短、抽穗扬花迟而不齐、叶片出现白绿条斑;
棉花缺锌叶片脉间失绿,边缘上卷,节间缩短,生育期推迟;烟草缺锌下部叶片的叶尖及叶缘出现水渍状失绿坏死斑点,叶小而厚,节间短;马铃薯缺锌株型矮缩,顶端叶片直立,叶小,叶面上出现灰色至古铜色的不规则斑点,叶缘上卷;
大豆缺锌叶片呈柠檬黄色并出现褐色斑点,逐渐扩大并连成坏死斑块;蚕豆缺锌出现“白苗”,成长后上部叶片变黄、叶形变小;
叶菜类蔬菜缺锌植株矮化,叶色发黄或铜青色有斑点;
番茄、青椒等果菜类缺锌小叶丛生状,新叶发生黄斑并逐渐向全叶扩展。
锰主要以Mn2+形式被植物吸收。锰是光合放氧复合体的主要成员,缺锰时光合放氧受到抑制。锰为形成叶绿素和维持叶绿素正常结构的必需元素。锰也是许多酶的活化剂,如一些转移磷酸的酶和三羧酸循环中的柠檬酸脱氢酶、草酰琥珀酸脱氢酶、α-酮戊二酸脱氢酶、苹果酸脱氢酶、柠檬酸合成酶等,都需锰的活化,故锰与光合和呼吸均有关系。锰还是硝酸还原的辅助因素,缺锰时硝酸就不能还原成氨,植物也就不能合成氨基酸和蛋白质。
缺锰时植物不能形成叶绿素,叶脉间失绿褪色,但叶脉仍保持绿色,此为缺锰与缺铁的主要区别。
钼以钼酸盐(MoO42-)的形式被植物吸收,当吸收的钼酸盐较多时,可与一种特殊的蛋白质结合而被贮存。
钼是硝酸还原酶的组成成分,缺钼则硝酸不能还原,呈现出缺氮病症。豆科植物根瘤菌的固氮特别需要钼,因为氮素固定是在固氮酶的作用下进行的,而固氮酶是由铁蛋白和铁钼蛋白组成的。
缺钼时叶较小,叶脉间失绿,有坏死斑点,且叶边缘焦枯,向内卷曲。十字花科植物缺钼时叶片卷曲畸形,老叶变厚且枯焦。禾谷类作物缺钼则籽粒皱缩或不能形成籽粒。
氯是在1954年才被确定的植物必需元素。氯以Cl-的形式被植物吸收。体内绝大部分的氯也以Cl-的形式存在,只有极少量的氯被结合进有机物,其中4氯吲哚乙酸是一种天然的生长素类激素。植物对氯的需要量很小,仅需几个mg•L-1,而盐生植物含氯相对较高,约70~100mg•L-1。
在光合作用中Cl-参加水的光解,叶和根细胞的分裂也需要Cl-的参与,Cl-还与K+等离子一起参与渗透势的调节,如与K+和苹果酸一起调节气孔开闭。
缺氯时,叶片萎蔫,失绿坏死,最后变为褐色;同时根系生长受阻、变粗,根尖变为棒状。
